Oxidative stress and the unfulfilled promises of antioxidant agents
Marco Giorgio
Department of Experimental Oncology, Institute of Oncology, Via Adamello 16, 20139, Milan, Italy
Correspondence to: Marco Giorgio. Email: marco.giorgio@ieo.eu
Abstract
It is well known that aging and its associated diseases, including cancer, are triggered by oxidative damage to biological macromolecules. However, antioxidant compounds are still disappointingly distant from any clinical application, so that Jim Watson has declared that antioxidant supplementation may have caused more cancers than it has prevented Watson J ((2013) Oxidants, antioxidants and the current incurability of metastatic cancers Open Biol 3 DOI: 10.1098/rsob.120144).
To clarify this paradox, here, we describe the mechanisms of oxidative stress focusing in particular on redox balance and physiological oxidative signals.
Keywords: oxidative stress, reactive oxygen species, mitochondrial respiration, redox signalling, antioxidant, cancer
Copyright: © the authors; licensee ecancermedicalscience. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Published: 23/07/2015; Received: 18/12/2014
What is oxidative stress?
Oxidative stress indicates a condition occurring when oxidising substances accumulate and accidental oxidative reactions thrive. In mammals, such as in all aerobic eukaryotes, the molecules with substantial oxidising potential contain oxygen. In particular, both reactive oxygen species (ROS), including singlet O2, superoxide anion (O2−), hydrogen peroxide (H2O2), and hydroxyl radical (OH−), and reactive nitrogen species (RNS), including peroxynitrite (ONOO−), nitrogen dioxide (NO2), and dinitrogen trioxide (N2O3), are potent oxidising agents in living organisms [2].
In cells, the accumulation of ROS and, subsequently, RNS increases the chance of formation of oxidative modifications in proteins, resulting in protein carbonyl content and oxidised- or nitro-modified residues, in lipids, generating hydroperoxide lipid derivatives, and in both purines and pyrimidines, inducing DNA adducts and breaks. Ultimately, the rise of oxidative attacks to biological macromolecules leads to the dysfunctions of proteins, membranes, and nucleic acids.
How do antioxidants work?
Antioxidants are substances that can neutralise ROS and RNS by accepting or donating electrons and, as a consequence, might be converted into a radical form containing unpaired electrons that are less reactive than the neutralised ROS or RNS. Then, another antioxidant molecule may regenerate the paired form of the radical antioxidant. Loosely held hydrogen, large aromatic structures, and unsaturated bonds are all potent electron acceptor sites able to neutralise the free radicals present in antioxidant molecules [3]. Several plant metabolites contain these antioxidant moieties, for example, the large class of polyphenol compounds, including quercitin, myricetin, catechins, anthocyanins, that are present in food sources [4].
Together with these plant derivatives, a variety of different antioxidants are available for clinical purposes. These include endogenous molecules, such as α-tocopherol, β-carotene, glutathione, ascorbic acid, adenosine, lipoic acid, coenzyme Q, lactoferrin, as well as synthetic antioxidants, such as thiols, ebselen (selenium-based peroxide scavenger) [5], idebenone (coenzyme Q-analogue) and the more recently developed mitoQ (mitochondrial-targeted ubiquinone) [6], or α-tocopheryl-succinate nanoparticles [7]. Notably, several cardioprotective drugs, such as probucol and carvediol, or neuroprotective drugs, such as edaravone, are potent and effective antioxidant against the overload of ROS upon ischemia/reperfusion [8].
Other compounds, often included in the category of antioxidants, increase instead the endogenous levels of endogenous antioxidants such as n-acetyl cysteine (precursor of glutathione) or inhibit ROS production from cellular oxidases (allopurinol) or from metal ion reactions as in the case of iron chelators (deferoxamine) [3].
Antioxidant failure in clinical studies
The present day is a golden age for antioxidants, promoted by the popularity of the free-radical theory of aging. Eating food rich in antioxidants protects from cancer and heart disease is common in particular.
Since the last three decades, the establishment of pre-clinical evidence showing that antioxidants protect deoxyribo nucleic acid (DNA) from being damaged by oxygen free radicals, potentially preventing the genetic mutations that cause cancer [9]. Antioxidants have also been consistently shown to reduce oxidative damage to low-density lipoprotein (LDL) cholesterol inside atherosclerotic plaques, thus protecting against atherosclerosis in the walls of arteries [10]. Thus, antioxidant treatments was promising to increase longevity by defeating the putative major cause of aging, that is, oxidative stress, or, more specifically by fighting top killers such as cancer and cardiovascular disease.
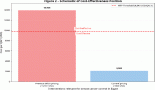
Unfortunately, several large randomised clinical trials found that antioxidant supplementation does not reduce the risk of cardiovascular disease or cancer, whereas one antioxidant, β-carotene, actually appears to increase the risk of some types of cancer in smokers. From 1985, the α-tocopherol/β-carotene cancer prevention trial (ATBC) was the first large study to examine the effect of antioxidants. Aged male smokers who had assumed 20 mg β-carotene for 8 years had an 18% increase in lung cancer incidence, and a less significant increase in prostate cancer, with respect to the placebo group [11]. In the following years, the beneficial effects of supplementation with β-carotene or vitamin A, α-tocopherol, ascorbic acid, and selenium increasingly became the object of debate [12–17].
Presently, after reviewing all the information, the US Preventive Services Task Force recommends against β-carotene or vitamin E to prevent cancer and cardiovascular disease (http://www.uspreventiveservicestaskforce.org) [18].
Furthermore, regardless of the fact that some epidemiological studies have overall shown that vegetable-enriched diets, while increasing antioxidant intake [19], inversely relate to mortality [20] or cancer [21] and stroke [22] risks, and that, in animal models, antioxidant-rich foods appears to inhibit tumorigenesis [23] and to be cardioprotective [24], both the US food and drug administration [25] and the European food safety authority [26] have banned any writing that could imply potential health benefits on the package labels of products with antioxidants.
Actually, the cancer prevention recommendations of eating, mostly, food of plant origin indicated by international health organisations such as the World Health Organisation (www.who.int/dietphysicalactivity/whatworks) or the World Cancer Research Fund International [27] are not related to vegetable antioxidant supply.
Insight into ROS metabolism
The negative effect of oxygen on living organisms has been known for a long time [28]. However, although a negative correlation was observed in vertebrates between the intracellular levels of ROS/oxidative stress and longevity, the levels of endogenous antioxidants were also found to anti-correlate with life span. The supplementation of antioxidants in animal models, including frogs, pigs, rats and mice, did not affect mortality [29]. In actual fact, oxidative stress is determined by the rates of both ROS production and scavenging. Thus, the negative effect of aerobic metabolism could be only partially balanced by antioxidant activities if ROS production is maintained.
ROS are usually considered as a side effect of aerobic metabolism, and mitochondrial respiration is thought to be the main intracellular source of accidental ROS [30]. During mitochondrial respiration, electrons are extracted from nicotinamide adenine dinucleotide (NADH) or succinate and are then transferred to O2 through a chain of enzymatic complexes. In the final step of this electron-transfer chain (ETC), the cytochrome c oxidase (complex IV) catalyses the full reduction of molecular O2 to water, without forming O2 radicals. However, partial reduction of O2 leading to the formation of O2− can occur if O2 hits sensible reduced sites of the ETC upstream of complex IV [31, 32]. Experimental data indicate that ROS are indeed continuously produced during mitochondrial respiration and that up to 2% of the total O2 consumption is converted to ROS [30].
Cells are normally able to defend themselves against ROS damage through the use of specific enzymatic (dismutases, catalase, peroxidases) or non-enzymatic (A, C, and E vitamins, uric acid, bilirubin) ROS-reducing mechanisms. The O2− dismutase enzyme, for instance, catalyses the conversion of O2− into H2O2, which is, in turn, reduced to water by the glutathione peroxidase and the catalase enzymes. In this way, the levels of different ROS are lowered to avoid the excessive oxidation of cellular components [30].
The rate of aging is assumed to be influenced, at least in part, by the rate of ROS production rather than by the rate of ROS scavenging. However, increased ROS production during aging is controversial; in contrast, scavenging activity has been clearly found to decrease over a lifetime and in different degenerative diseases [33–35].
Oxidative stress has a physiological role
Substantial evidence demonstrates that ROS have a physiological role regardless of their toxicity. In fact, ROS have been shown to mediate growth factor/hormone/cytokine signal transduction, to regulate gene expression, and to determine programmed cell death [36, 37]. Among ROS, H2O2 is diffusible, less reactive and longer-lived than, for instance, O2− and OH−. H2O2 is especially involved in the regulation of intracellular signalling pathways and could be considered as a second messenger [37].
Since the rate of mitochondrial ROS formation depends on the local concentrations of O2 and energetic substrates, and on ATP cellular demands [38, 39], the emerging picture is that mitochondria generate ROS in a regulated manner to deal with the different metabolic activities of the cell [40] and/or hypoxic conditions [41, 42].
ROS do not only cause irreversible damage to cellular components, they can also lead to fully reversible protein modifications. In particular, H2O2 has been demonstrated to directly oxidise cysteinyl thiols inducing formation of disulphide bonds and sulphenic acids. It has also been shown to induce glutathionylation of cysteine residues or the formation of methionine sulphoxide on methionine residues in a variety of contexts, such as the transcription factors OxyR and Pap1 in bacteria and in the yeast, respectively, the Kinase Sty1 in the yeast, the vacuolar ATPase, Vatp, in plants, the HIV-2 protease in viruses, the arylamine N-acetyl transferase 1, NAT1, the indoleamine 2, 3-dioxigenase, the phospholipase A2, iPLA2Beta, the small ubiquitin-related modifiers SUMO E1 subunit Uba2 and SUMO E2-conjugating enzymes Ubc9, the phosphatases PTP1B (protein tyrosine phosphatase 1B) and PTEN (phosphatase and tensin homologue), the peroxidase enzymes Prx I and II (both only cytosolic) and Prx III (cytosolic and mitochondrial), the annexin A2 protein, the heat shock factor 1 (HSF1) the mitochondrial enzymes aconitase and α-ketoglutarate dehydrogenase, and the subunits of the respiratory complex I, in mammals [43].
Overall, it has been demonstrated that oxidation and reduction of key cellular proteins participate in a redox-dependent regulation of cellular functions, including energy metabolism and response to stress. It has also become clear that intracellular signalling pathways can be activated by changes in intracellular metabolic redox reactions that involve O2− and H2O2 [44]. Early hypotheses had proposed that exposure to specific environmental factors can induce ROS accumulation, thus triggering abnormal ROS-signalling leading to increase proliferation and malignant transformation. Evidence for this was obtained from studies showing that carcinogen initiators and promoters, including ionising radiation and polycyclic aromatic hydrocarbons, increase ROS formation, which in turn favours tumorigenesis [45]. A clear mechanism of how H2O2 in particular, can favour proliferation has emerged from studies on the redox regulation of critical phosphatases involved in signal transduction from plasma membrane receptors, together with the findings that several growth factors, such as EGF or Insulin/IGF, trigger H2O2 production directly from their membrane receptors [51, 79].
In this context, the function of the p66Shc protein is representative. P66Shc is the largest of the three isoforms encoded by the ShcA locus and almost ubiquitously expressed in vertebrates; it functions to regulate intracellular ROS levels and mitochondrial apoptosis. Cytosolic p66Shc mediates activation of the membrane oxidase activity and suppresses catalase and MnSOD expressions [46]. Then, a fraction of p66Shc translocates within the mitochondrial inter-membrane space [47] upon specific stimuli, including pro-apoptotic stresses [48] or growth factor stimulation [49], and oxidises cytochrome c to form H2O2 [50], which in turn regulates mitochondrial [51], and cellular functions [49, 52]. Accordingly, cells from p66Shc null mice or p66Shc-depleted by RNAi have reduced ROS levels [49]; however, p66Shc null mice show normal tumour incidence [53] and increased mutation rate [54]. Notably, p66Shc deletion is counter-selected when mice are maintained in harsh settings that mimic conditions in the wild (in an open field in the cold and in competition for food), indicating that the pro-oxidant function of p66Shc is essential for fitness under stressing natural conditions but redundant in a protected environment [55].
The role of oxidative stress in cancer
Tumorigenesis is characterised by major alterations in energetic metabolism, O2 consumption and ROS accumulation [2] which result in a change in the balance between reduced/oxidised species (redox balance) [56]. Changes in the cellular redox balance affect proliferation, migration, and survival of cancer cells contributing to disease progression [57–59]. Activated oncogenes, such as Myc [60], Bcl-2 [61] or Ras [62], have been reported to affect redox balance. For example, the expression of the oncogenic form of Ras was found to boost [63–65] or to reduce [66] the level of glutathione, depending on the cell line. Furthermore, a reducing environment associates with different types of cancer [67–68]. This “reducing” environment results from the relative concentration of all the oxidant and reducing species (redox species) that exist in the metabolic network [69]. As a consequence, high levels of ROS endogenous scavengers or treatment with antioxidants increase oncogenic transformation [70] or tumour progression [71, 72], whereas increasing oxidation has even been proposed as a therapeutic strategy for cancer [73–76]. Finally, the antioxidant activity of chemicals is considered hazardous for novel classes of anticancer drugs [77, 78].
Conclusion
Antioxidants affecting ROS levels and functions produce different outcomes since ROS have both deleterious and beneficial effects. Although ROS are a by-product of aerobic metabolism, several enzymatic systems have evolved to generate ROS on purpose. In cancer cells, ROS act as secondary messengers of oncogenic signalling pathways and can also induce cellular senescence and apoptosis. As a consequence, oxidative stress during cancer expansion and progression selects for clones with high antioxidant metabolism. Based on this, the failure of antioxidant treatments, as documented in several clinical trials, is not surprising. Antioxidant drugs are not sufficient to inhibit tumorigenesis or, even worst, they may accelerate it.
References
1. Watson J (2013) Oxidants, antioxidants and the current incurability of metastatic cancers Open Biol 3 DOI: 10.1098/rsob.120144
2. Halliwell B and Gutteridge JMC (2007) Free Radicals in Biology and Medicine 4th edn Oxford University Press Oxford
3. Cadenas E and Packer L (2002) Hand Book of Antioxidants (New York: Marcel Dekker Inc) Introduction, Chapters 1, 6
4. Halliwell B (2002) Food Derived Antioxidant: How to Evaluate Their Importance in Food and In Vivo In: Cadenas E, Packer L (ed) Handbook of antioxidants (New York: Marcel Inc) Chapter 1.1
5. Fortmann SP, Burda BU, Senger CA, Lin JS and Whitlock EP (2013) Vitamin and mineral supplements in the primary prevention of cardiovascular disease and cancer: an updated systematic evidence review for the U.S. preventive services task force Ann Intern Med 159 824–834 PMID: 24217421
6. Kelso GF, Porteous CM, Coulter CV and Hughes G et al (2001) Selective targeting of a redox-active ubiquinone to mitochondria within cells: antioxidant and antiapoptotic properties J Biol Chem 276 4588–4596 DOI: 10.1074/jbc.M009093200
7. Hama S and Kogure K (2014) Nanoparticles consisting of tocopheryl succinate are a novel drug-delivery system with multifaceted antitumor activity Biol Pharm Bull 37 196–200 DOI: 10.1248/bpb.b13-00848 PMID: 24492715
8. Lefer DJ and Granger DN (2000) Oxidative stress and cardiac disease Am J Med 109 315–323 DOI: 10.1016/S0002-9343(00)00467-8 PMID: 10996583
9. Ames BN (1983) Dietary carcinogens and anticarcinogens Oxygen radicals and degenerative diseases Science 221 1256–1264 DOI: 10.1126/science.6351251 PMID: 6351251
10. Morel DW, DiCorleto PE and Chisolm GM (1984) Endothelial and smooth muscle cells alter low density lipoprotein in vitro by free radical oxidation Arteriosclerosis 4 357–364 DOI: 10.1161/01.ATV.4.4.357 PMID: 6466193
11. The Alpha-Tocopherol, Beta Carotene Cancer Prevention Study Group (1994) The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers N Engl J Med 330 1029–1235 DOI: 10.1056/NEJM199404143301501
12. Lawlor DA, Smith GD, Bruckdorfer KR, Kundu D and Ebrahim S (2004) Those confounded vitamins: what can we learn from the differences between observational versus randomised trial evidence? Lancet 363 1724–1727 DOI: 10.1016/S0140-6736(04)16260-0 PMID: 15158637
13. Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG and Gludd C (2007) Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: systematic review and meta-analysis JAMA 297 842–857 DOI: 10.1001/jama.297.8.842 PMID: 17327526
14. Greenlee H, Hershman DL and Jacobson JS (2009) Use of antioxidant supplements during breast cancer treatment: a comprehensive review Breast Cancer Res Treat 115 437–452 DOI: 10.1007/s10549-008-0193-0
15. Lippman SM et al (2009) Effect of selenium and vitamin E on risk of prostate cancer and other cancers: the selenium and vitamin E cancer prevention trial (SELECT) JAMA 301 39–51 DOI: 10.1001/jama.2008.864
16. Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG and Gluud C (2012) Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases Cochrane Database Syst Rev 3 CD007176 PMID: 22419320
17. Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG and Gluud C (2013) Meta-regression analyses, meta-analyses, and trial sequential analyses of the effects of supplementation with beta-carotene, vitamin A, and vitamin E singly or in different combinations on all-cause mortality: do we have evidence for lack of harm? PLoS One 8 e74558 DOI: 10.1371/journal.pone.0074558 PMID: 24040282 PMCID: 3765487
18. Coulter I, Hardy M and Shekelle P et al (2003) Effect of the Supplemental Use of Antioxidants Vitamin C, Vitamin E, and Coenzyme Q10 for the Prevention and Treatment of Cancer Evidence Report/Technology Assessment Number 75. (Prepared by Southern California Evidence-based Practice Center under Contract No. 290-97-0001) AHRQ Publication No. 04-E003 (Rockville, MD: Agency for Healthcare Research and Quality)
19. Park SY, Ollberding NJ and Woolcott CG et al (2013) Fruit and vegetable intakes are associated with lower risk of bladder cancer among women in the multiethnic cohort study J Nutr 143 1283–1292 DOI: 10.3945/jn.113.174920 PMID: 23739308 PMCID: 3709993
20. Masala G, Assedi M and Bendinelli B et al Fruit and vegetables consumption and breast cancer risk: the EPIC Italy study Breast Cancer Res Treat 132 1127–1136 PMID: 22215387
21. Genkinger JM, Platz EA, Hoffman SC, Comstock GW and Helzlsouer KJ (2004) Fruit, vegetable, and antioxidant intake and all-cause, cancer, and cardiovascular disease mortality in a community-dwelling population in Washington County, Maryland Am J Epidemiol 160 1223–1233 DOI: 10.1093/aje/kwh339 PMID: 15583375
22. Wang X, Ouyang Y and Liu J et al (2014) Fruit and vegetable consumption and mortality from all causes, cardiovascular disease, and cancer: systematic review and dose-response meta-analysis of prospective cohort studies BMJ 349 g4490 DOI: 10.1136/bmj.g4490 PMID: 25073782 PMCID: 4115152
23. Butelli E, Titta L and Giorgio M et al (2008) Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors Nat Biotechnol 26 1301–1308 DOI: 10.1038/nbt.1506 PMID: 18953354
24. Testai L, Martelli A and Cristofaro M et al (2013) Cardioprotective effects of different flavonoids against myocardial ischaemia/reperfusion injury in Langendorff-perfused rat hearts J Pharm Pharmacol 65 750–756 DOI: 10.1111/jphp.12032 PMID: 23600393
25. Guidance for industry, food labeling; nutrient content claims; definition for “high potency” and definition for “antioxidant” for use in nutrient content claims for dietary supplements and conventional foods U.S. department of health and human services, food and drug administration Center for food safety and applied nutrition June 2008
26. Scientific opinion on the substantiation of health claims related to various food(s)/food constituent(s) and protection of cells from premature aging, antioxidant activity, antioxidant content and antioxidant properties, and protection of DNA, proteins and lipids from oxidative damage pursuant to Article 13(1) of Regulation (EC) No 1924/20061 EFSA panel on dietetic products, nutrition and allergies (NDA)2, 3 European food safety authority (EFSA), Parma, Italy, EFSA J 2010 8 1489
27. World Cancer Research Fund/American Institute for Cancer Research (2007) Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective Washington DC: AICR
28. Lorrain SJ (1899) The pathological effects due to increase of oxygen tension in the air breathed J Physiol 24 19 DOI: 10.1113/jphysiol.1899.sp000746
29. Barja G (2004) Aging in vertebrates, and the effect of caloric restriction: a mitochondrial free radical production-DNA damage mechanism? Biol Rev Camb Philos Soc 79 235–251 DOI: 10.1017/S1464793103006213 PMID: 15191224
30. Chance B, Sies H and Boveris A (1979) Hydroperoxide metabolism in mammalian organs Physiol Rev 59 527–605 PMID: 37532
31. Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335–344 DOI: 10.1113/jphysiol.2003.049478 PMID: 14561818 PMCID: 2343396
32. Brand MD (2010) The sites and topology of mitochondrial superoxide production Exp Gerontol 45 466–472 DOI: 10.1016/j.exger.2010.01.003 PMID: 20064600 PMCID: 2879443
33. Frisard M and Ravussinm E (2006) Energy metabolism and oxidative stress: impact on the metabolic syndrome and the aging process Endocrine 29 27–32 DOI: 10.1385/ENDO:29:1:27 PMID: 16622290
34. Harman D (1998) Aging: phenomena and theories Ann N Y Acad Sci 854 1–7 DOI: 10.1111/j.1749-6632.1998.tb09886.x
35. Balaban RS, Nemoto S and Finkel T (2005) Mitochondria, oxidants, and aging Cell 120 483–495 DOI: 10.1016/j.cell.2005.02.001 PMID: 15734681
36. Orrenius S, Gogvadze V and Zhivotovsky B Mitochondrial oxidative stress: implications for cell death Annu Rev Pharmacol Toxicol 47(2007) 143–183 PMID: 17029566
37. Stone JR and Yang S (2006) Hydrogen peroxide: a signaling messenger Antioxid Redox Signal 8 243–270 DOI: 10.1089/ars.2006.8.243 PMID: 16677071
38. Hoffman DL and Brookes PSJ (2009) Oxygen sensitivity of mitochondrial reactive oxygen species generation depends on metabolic conditions Biol Chem 284 16236–16245 DOI: 10.1074/jbc.M809512200
39. Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1–13 DOI: 10.1042/BJ20081386
40. Marcu R, Rapino S and Trinei M et al (2012) Electrochemical study of hydrogen peroxide formation in isolated mitochondria Bioelectrochemistry 85 21–28 DOI: 10.1016/j.bioelechem.2011.11.005
41. Hoffman DL, Salter JD and Brookes PS (2007) Response of mitochondrial reactive oxygen species generation to steady-state oxygen tension: implications for hypoxic cell signaling Am J Physiol Heart Circ Physiol 292 H101–H108 DOI: 10.1152/ajpheart.00699.2006
42. Bell EL, Klimova TA and Eisenbart J et al (2007) The Qo site of the mitochondrial complex III is required for the transduction of hypoxic signaling via reactive oxygen species production Cell Biol 177 1029–1036 DOI: 10.1083/jcb.200609074
43. Giorgio M, Trinei M and Migliaccio E et al (2007) Hydrogen peroxide: a metabolic by-product or a common mediator of ageing signals? Nat Rev Mol Cell Biol 8 722–728 DOI: 10.1038/nrm2240 PMID: 17700625
44. Valko M, Leibfritz D and Moncol J et al (2007) Free radicals and antioxidants in normal physiological functions and human disease Int J Biochem Cell Biol 39 44–84 DOI: 10.1016/j.biocel.2006.07.001
45. Gius D and Spitz DR (2006) Redox signaling in cancer biology Antioxid Redox Signal 8 1249–1252 DOI: 10.1089/ars.2006.8.1249 PMID: 16910772
46. Trinei M, Berniakovich I and Beltrami E et al (2009) P66Shc signals to age Giorgio Aging 1 503–585
47. Orsini F, Moroni M and Contursi C et al (2006) Regulatory effects of the mitochondrial energetic status on mitochondrial p66Shc Biol Chem 387 1405–1410 DOI: 10.1515/BC.2006.176 PMID: 17081113
48. Pinton P, Rimessi A and Marchi S et al (2007) Protein kinase C beta and prolyl isomerase 1 regulate mitochondrial effects of the life-span determinant p66Shc Science 315 659–663 DOI: 10.1126/science.1135380 PMID: 17272725
49. Berniakovich I, Trinei M and Stendardo M et al (2008) P66Shc-generated oxidative signal promotes fat accumulation J Biol Chem 283 34283–34293 DOI: 10.1074/jbc.M804362200 PMID: 18838380 PMCID: 2590696
50. Trinei M, Migliaccio E and Bernardi P (2013) P66Shc, mitochondria, and the generation of reactive oxygen species Methods Enzymol 528 99–110 DOI: 10.1016/B978-0-12-405881-1.00006-9 PMID: 23849861
51. Trinei M, Berniakovich I and Pelicci PG et al (2006) Mitochondrial DNA copy number is regulated by cellular proliferation: a role for Ras and p66(Shc) Biochim Biophys Acta 1757 624–630 DOI: 10.1016/j.bbabio.2006.05.029 PMID: 16829231
52. Frijhoff J, Dagnell M and Augsten M et al (2014) The mitochondrial reactive oxygen species regulator p66Shc controls PDGF-induced signaling and migration through protein tyrosine phosphatase oxidation Free Radic Biol Med 68 268–277 DOI: 10.1016/j.freeradbiomed.2013.12.022 PMID: 24378437
53. Beltrami E, Valtorta S and Moresco R et al (2013) The p53-p66Shc apoptotic pathway is dispensable for tumor suppression whereas the p66Shc-generated oxidative stress initiates tumorigenesis Curr Pharm Des 19 2708–2714 DOI: 10.2174/1381612811319150005
54. Beltrami E, Ruggiero A and Busuttil R et al (2013) Deletion of p66Shc in mice increases the frequency of size-change mutations in the lacZ transgene Aging Cell 12 177–183 DOI: 10.1111/acel.12036
55. Giorgio M, Berry A and Berniakovich I et al (2012) The p66(Shc) knocked out mice are short lived under natural condition Aging Cell 11 162–168 DOI: 10.1111/j.1474-9726.2011.00770.x
56. Gupta SC, Hevia D and Patchva S et al (2012) Upsides and downsides of reactive oxygen species for cancer: the roles of reactive oxygen species in tumorigenesis, prevention, and therapy Antioxid Redox Signal 16 1295–1322 DOI: 10.1089/ars.2011.4414 PMCID: 3324815
57. Burhans WC and Heintz NH (2009) The cell cycle is a redox cycle: linking phase-specific targets to cell fate Free Radic Biol Med 47 1282–1293 DOI: 10.1016/j.freeradbiomed.2009.05.026 PMID: 19486941
58. Hitchler MJ and Domann FE (2012) Redox regulation of the epigenetic landscape in cancer: a role for metabolic reprogramming in remodeling the epigenome Free Radic Biol Med 53 2178–2187 DOI: 10.1016/j.freeradbiomed.2012.09.028 PMID: 23022407 PMCID: 3508253
59. Ward PS and Thompson CB (2012) Metabolic reprogramming: a cancer hallmark even warburg did not anticipate Cancer Cell 21 297–308 DOI: 10.1016/j.ccr.2012.02.014 PMID: 22439925 PMCID: 3311998
60. Egler RA, Fernandes E and Rothermund K et al (2005) Regulation of reactive oxygen species, DNA damage, and c-Myc function by peroxiredoxin 1 Oncogene 24 8038–8050 DOI: 10.1038/sj.onc.1208821 PMID: 16170382
61. Low IC, Kang J and Pervaiz S (2011) Bcl-2: a prime regulator of mitochondrial redox metabolism in cancer cells Antioxid Redox Signal 15 2975–2987 DOI: 10.1089/ars.2010.3851 PMID: 21574773
62. Armeni T, Ercolani L and Urbanelli L (2012) Cellular redox imbalance and changes of protein S-glutathionylation patterns are associated with senescence induced by oncogenic H-ras PLoS One 7 e52151 DOI: 10.1371/journal.pone.0052151
63. Chuang JIM, Chang TY and Liu HS (2003) Glutathione depletion-induced apoptosis of Ha-ras-transformed NIH3T3 cells can be prevented by melatonin Oncogene 22 1349–1357 DOI: 10.1038/sj.onc.1206289 PMID: 12618760
64. Recktenwald CV, Kellner R and Lichtenfels R (2008) Altered detoxification status and increased resistance to oxidative stress by K-ras transformation Cancer Res 68 10086–10093 DOI: 10.1158/0008-5472.CAN-08-0360 PMID: 19074874
65. Vincenzini MT, Marraccini P and Iantomasi T et al (1993) Altered metabolism of glutathione in cells transformed by oncogenes which cause resistance to ionizing radiations FEBS Lett 320 219–223 DOI: 10.1016/0014-5793(93)80590-Q PMID: 8096467
66. Choudhary S, Rathore K and Wang HC (2011) Differential induction of reactive oxygen species through Erk1/2 and Nox-1 by FK228 for selective apoptosis of oncogenic H-Ras-expressing human urinary bladder cancer J82 cells J Cancer Res Clin Oncol 137 471–480 DOI: 10.1007/s00432-010-0910-z
67. Schumacker PT (2006) Reactive oxygen species in cancer cells: live by the sword, die by the sword Cancer Cell 10 175–176 DOI: 10.1016/j.ccr.2006.08.015 PMID: 16959608
68. Smart DK, Ortiz KL and Mattson D et al (2004) Thioredoxin reductase as a potential molecular target for anticancer agents that induce oxidative stress Cancer Res 64 6716–6724 DOI: 10.1158/0008-5472.CAN-03-3990 PMID: 15374989
69. Schafer FQ and Buettner GR (2001) Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple Free Radic Biol Med 30 1191–1212 DOI: 10.1016/S0891-5849(01)00480-4 PMID: 11368918
70. DeNicola GM et al (2011) Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis Nature 475 106–109 DOI: 10.1038/nature10189 PMID: 21734707 PMCID: 3404470
71. Schafer ST et al (2009) Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment Nature 461 109–113 DOI: 10.1038/nature08268 PMID: 19693011 PMCID: 2931797
72. Sayin VI, Ibrahim MX and Larsson E et al (2014) Antioxidants accelerate lung cancer progression in mice Sci Transl Med 6 221ra15 DOI: 10.1126/scitranslmed.3007653 PMID: 24477002
73. Montero AJ and Jassem J (2011) Cellular redox pathways as a therapeutic target in the treatment of cancer Drugs 71 1385–1396 DOI: 10.2165/11592590-000000000-00000 PMID: 21812504
74. Gorrini C, Harris IS and Mak TW (2013) Modulation of oxidative stress as an anticancer strategy Nat Rev Drug Discov 12 931–947 DOI: 10.1038/nrd4002 PMID: 24287781
75. Glasauer A, Sena LA and Diebold LP et al (2014) Targeting SOD1 reduces experimental non-small-cell lung cancer J Clin Invest 124 117–128 DOI: 10.1172/JCI71714 PMCID: 3871252
76. Sullivan LB and Chandel NS (2014) Mitochondrial reactive oxygen species and cancer Cancer Metab 2 17 DOI: 10.1186/2049-3002-2-17
77. Neuzil J, Tomasetti M and Zhao Y et al (2007) Vitamin E analogs, a novel group of “mitocans,” as anticancer agents: the importance of being redox-silent Mol Pharmacol 71 1185–1199 DOI: 10.1124/mol.106.030122 PMID: 17220355
78. Lawenda BD, Kelly KM and Ladas EJ et al (2008) Should supplemental antioxidant administration be avoided during chemotherapy and radiation therapy? J Natl Cancer Inst 100 773–783 DOI: 10.1093/jnci/djn148 PMID: 18505970
79. Heneberg P (2014) Reactive nitrogen species and hydrogen sulfide as regulators of protein tyrosine phosphatase activity Antioxid Redox Signal 29 2191-209 DOI: 10.1089/ars.2013.5493 PMID: 24328688